脂滴(Lipid Droplets,LDs)具有磷脂单层,围绕由疏水性中性脂质组成的核心。脂滴由内质网产生,表面分布着一组特定的蛋白质,这些蛋白质通过疏水发夹(图1a左)、脂质化结构域和两亲性螺旋或与脂滴表面的另一种蛋白质结合(图1a右)而与单层相关联。在出芽酵母中,脂滴的平均直径为200 nm,而在脂肪细胞分化的早期和晚期,脂滴直径分别约为10和100 mm。
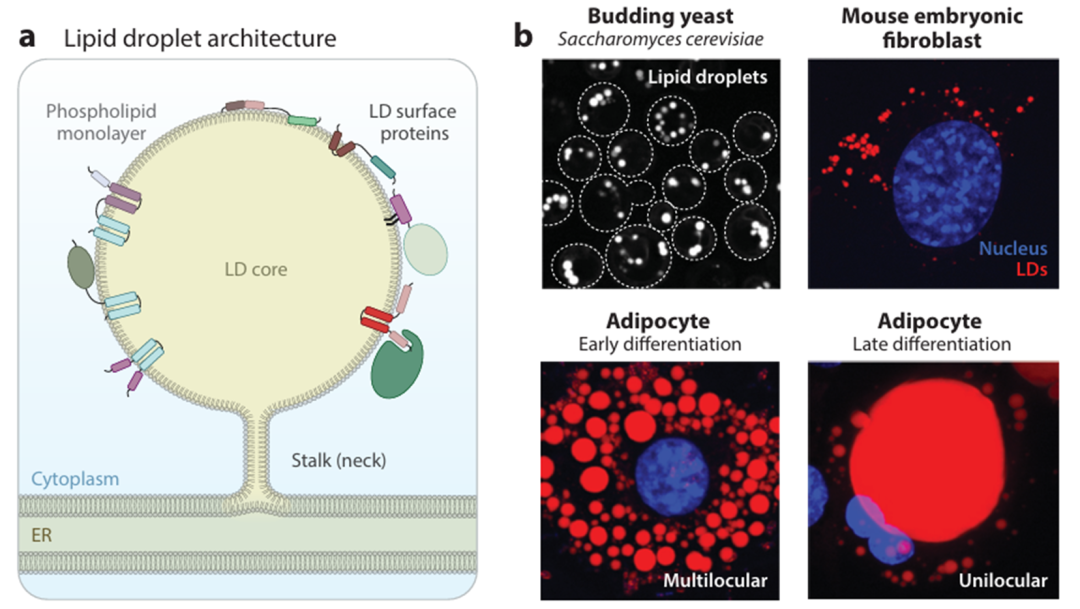
图1 (a)LDs基本结构示意图。(b)通过亲脂性染料(BODIPY)可视化静止出芽酵母细胞中的脂滴分布(虚线表示细胞轮廓)。使用LipidTOX red对胚胎成纤维细胞中的脂滴进行染色[核(蓝色)]。3T3-L1前脂肪细胞分化为脂肪细胞6天(早期分化,多室脂滴)或4周(晚期分化,单室脂滴)。多室脂滴和单室脂滴用LipidTOX red进行可视化。请注意,图像未按比例显示。
脂类(lipids),又称脂质,包括脂肪和类脂,其共同属性是具有脂溶性。脂肪即三酰甘油(也称甘油三酯)(triacylglycerol,TAG),类脂主要指胆固醇和磷脂。TAG由一个甘油分子和三个脂肪酸形成的酯;人体内含量最多的磷脂是甘油磷脂,即三酰甘油中一个或两个脂肪酸被含磷酸的基团取代的脂类。根据X取代基的不同,甘油磷脂又可分为多种:取代基是胆碱的为卵磷脂,是乙醇胺的为脑磷脂,是丝氨酸的为丝氨酸磷脂。

脂肪酸(fattyacides, FAs)由碳、氢、氧三种元素组成。碳链中不含双键的脂肪酸为饱和脂肪酸(SFA),含有双键的脂肪酸为不饱和脂肪酸。依据碳链中含双键的数目将不饱和脂肪酸分为:① 单不饱和脂肪酸(MUFA),碳链中只含一个双键;② 多不饱和脂肪酸(PUFA),碳链中含两个以上双键。在大多数细胞中,代谢能量以脂肪酸(FAs)的形式储存。根据碳链上第一个双键的位置,不饱和脂肪酸可分为(ω-3、 ω-6等)。直链脂肪酸中距离羧基最远的碳原子称ω碳原子,若从ω 碳原子起(即从甲基端数起)第一个双键在第三和第四碳原子之间的不饱和脂肪酸,称为ω-3系列脂肪酸,以此类推。
图2 EPA和DHA的分子式
1. 脂滴的发生
细胞质脂滴发生涉及合成、成核、胞质出芽和生长(图3①)。
(
1
)成核:三酰甘油(
TAGs
)在内质网(
ER
)膜中积累,并在达到临界凝聚浓度(约
5–10 mol%
)以上时发生相分离,形成透镜结构,随后被
BSCL2-Seipin
复合物结合。(
2
)出芽:在
LDAF
(后生动物)和
Ldo45
(酵母)的帮助下,
LDs
向细胞质中出芽。出芽的方向性取决于多种因素,包括
FIT2
。
I
类和
ERTOLD
蛋白可以在此阶段从内质网(
ER
)进入
LD
表面。
(3
)早期扩张:三酰甘油(
TAGs
)的持续增加导致
LD
尺寸扩大。其表面通过
Seipin
复合物(
ERTOLD
途径)与内质网(
ER
)形成的连接,增加磷脂和
I
类蛋白,以及直接从细胞质(
CYTOLD
途径)中添加
II
类蛋白和伴侣蛋白辅助的蛋白来生长。(
4
)晚期扩张:一些脂质滴(
LD
)表面蛋白(例如,
GPAT4
和
ATGL
)被认为在扩张后期才到达
LD
,它们并非通过
Seipin
进入表面,而是通过在内质网(
ER
)和脂质滴(
LD
)之间形成的桥进入。
TAGs
的生物合成主要通过“甘油
-3-
磷酸途径”完成:
脂肪酸(
FAs
)与辅酶
A
(
CoA
)结合并被激活,随后与甘油
-3-
磷酸(
G-3-P
)酯化,生成溶血磷脂酸(
LPA
)和磷脂酸(
PA
)。磷脂酸(
PA
)去磷酸化生成二酰甘油(
DAG
),后者继而与第三个脂肪酸(
FA
)酯化,生成三酰甘油(
TAG
)。
脂滴不仅是储存单位,也是代谢单位。脂滴的分解代谢有两个主要机制:脂解(lipolysis)和脂肪吞噬(lipophagy)。
Zadoorian A, Du X, Yang H. Lipid droplet biogenesis and functions in health and disease. Nat Rev Endocrinol. 2023 Aug;19(8):443-459. doi: 10.1038/s41574-023-00845-0. Epub 2023 May 23. PMID: 37221402; PMCID: PMC10204695.
Klemm RW, Carvalho P. Lipid Droplets Big and Small: Basic Mechanisms That Make Them All. Annu Rev Cell Dev Biol. 2024 Oct;40(1):143-168. doi: 10.1146/annurev-cellbio-012624-031419. PMID: 39356808.
脂滴可视化主要包括下述两种:
(1)基于脂滴包被蛋白(perilipin,PLIN)家族蛋白的免疫荧光定位;
(2)荧光探针。传统染料,如苏丹红III和油红O已用于对 LD 进行染色,但已发现它们在染色过程中会破坏 LD 结构。还可通过荧光染料,如尼罗河红、BODIPY和 LipidTox。
5. 脂滴与线粒体
脂滴包被蛋白(perilipin,PLIN)5促进脂肪酸(FA)从脂滴流向线粒体,并在细胞器之间提供物理连接;其同源物PLIN1在脂肪细胞中特异性表达,已被证明与线粒体融合蛋白2 (Mfn2)直接相互作用,而Mfn2是线粒体融合和线粒体-脂滴相互作用的介质。
在骨骼肌中,小G蛋白Rab8a与PLIN5相互作用,调控脂滴与线粒体的接触,并响应AMPK的激活;而饥饿状态下,脂酰辅酶A合成酶FATP4与PLIN5相互作用,增加脂滴与线粒体的接触,二者都能够促进脂肪酸从脂滴转移到线粒体进行β-氧化。
铁(Fe2+)通过与过氧化氢(H2O2)反应被氧化成铁(Fe3+),从而形成高活性的羟基自由基(OH·),称为芬顿反应。Fe3+通过与超氧自由基(O2
·-
)的反应被还原成Fe2+。这个氧化还原循环被称为HarberWeiss反应。在非酶性脂质过氧化中,自由基,如羟基自由基(
OH·
),从多不饱和脂肪酸(
PUFA
)中抽出一个氢,形成一个碳心磷脂(
PL
)自由基(
PL·
)。
PL.
与分子氧(
O2
)反应,形成磷脂过氧自由基(
PLOO·
)。
PLOO.
从另一个
PUFA
中提取氢,形成磷脂过氧化氢(
PLOOH
)和一个新的
PL.
,它可以再次与
O2
反应。在酶法脂质过氧化中,脂氧酶(
LOX
)催化
PUFAs
的二氧化并产生
PLOOH
。一方面,
PLOOH
在亚铁(
Fe2+
)的存在下可以分解成烷氧基磷脂自由基(
PLO·
),通过攻击另一种
PUFA
促进脂质过氧化的进一步传播。另一方面,
PLOOH
可能分解为
4-
羟基壬烯醛(
4-HNE
)或丙二醛(
MDA
),通过交联可能使蛋白质失活。
PLs
的过氧化和
4-HNE
或
MDA
的产生导致膜不稳定和通透性,从而导致细胞死亡。谷胱甘肽过氧化物酶
4
(
GPX4
)具有独特的能力,可以将活性
PLOOH
还原为非活性
PL
醇(
PL-OH
),从而中断自由基连锁反应,抑制脂质过氧化。
(1)SystemXc-调控:System Xc-由SLC3A2和SLC7A11二聚体组成,嵌于细胞膜表面。SLC7A11是发挥功能的主要亚基,可将胱氨酸转运入胞,用于合成GSH;因此,抑制SLC7A11表达可诱导铁死亡发生。研究发现,在人肿瘤细胞中SLC7A11过度表达,这种过表达能够抑制活性氧诱导的“铁死亡”,同时削弱p533KR介导的对肿瘤生长的抑制作用。
(2)GSH和GPX4调控:研究认为GPX4是细胞生存的关键,也是铁死亡的核心调控蛋白,GPX4能降解小分子过氧化物和某些脂质过氧化物,抑制脂质过氧化。果不能有效地被Gpx4淬灭,磷脂氢过氧化物能够在过渡金属(如铁)存在的情况下引发催化反应,最终导致细胞死亡。研究发现,若细胞中GPX4 表达下调则会对铁死亡更敏感;敲降GPX4即可诱导铁死亡发生;相反,若上调GPX4 的表达,则会产生对铁死亡的耐受。
(3)ROS调控:诱导铁死亡发生均会导致细胞内脂质ROS升高,从而引起细胞死亡,这也是脂质抗氧化剂可抑制铁死亡的原因。线粒体作为含铁丰富(铁是线粒体氧化呼吸链必需离子)、ROS产生为主的细胞器,被认为是铁死亡发生的重要场所。它既是细胞内产生ROS的重要场所,其脂肪酸代谢又为细胞铁死亡提供特定的脂质前体。p53也能够通过ROS应答和"铁死亡"抑制肿瘤生长。
酸性条件下,PUFA通过促进脂质过氧化,诱导细胞的铁死亡,而脂滴可以吸收PUFA而抵抗酸性环境的细胞毒性;
当酸性癌细胞不能缓冲PUFAs的高水平吸收,并暴露在过氧化的有害影响下时,会促进铁死亡发生;
抑制脂滴的合成有利于PUFA诱导肿瘤的铁死亡;
小鼠肿瘤模型中,高ω-3 PUFA饮食可以增强铁死亡诱导剂对肿瘤的杀伤力。

